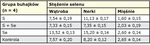
Dość interesująca jest współzależność siarki i selenu (Se), które należą do tej samej grupy okresowego układu pierwiastków i wykazują podobne właściwości chemiczne. Dlatego często ze sobą konkurują i wzajemnie się zastępują (6), ale także mogą ze sobą reagować, tworząc nierozpuszczalne siarczki, i razem wchodzić w skład selenowych pochodnych aminokwasów siarkowych, polipeptydów i białek. Często też obydwa pierwiastki występują razem w dużej ilości w kopalinach, m.in. w węglu i ropie, a w efekcie tego także w środowisku zwierząt. Wspólne występowanie, a zarazem konkurencyjność wobec siebie mogą być korzystne w przypadku równoczesnego ich nadmiaru i wzajemnej neutralizacji, ale także niekorzystne, gdy jest nadmiar S i niedobór Se. Nadmiar S w glebie i następowy wzrost jej kwasowości może ograniczać przyswajalność selenu przez rośliny, a tym samym zmniejszać jego pobór przez zwierzęta roślinożerne. Z kolei następstwem hiposelenozy u tych zwierząt jest zwyrodnienie mięśni i ciężkie objawy kliniczne, określane terminem pokarmowej dystrofii mięśni. Masowe wystąpienie tej choroby obserwowano u źrebiąt na terenach bogatych w S i o kwaśnym odczynie gleb oraz o niskiej zawartości Se w miejscowej roślinności (10). Na możliwość negatywnego wpływu nadmiaru S na przyswajalność i metabolizm Se wskazują też wyniki badań doświadczalnych. Obserwowano zwykle, choć nie zawsze, spadek zawartości Se w surowicy i w wątrobie oraz zwiększone jego wydalanie z moczem (6, 7, 17). Nie potwierdzają tego w pełni wyniki badań Riha (28), których odnośną część przedstawia tab. III i IV.
Dodatek siarki elementarnej nie wpływał na stężenie Se w surowicy, ale dodatek seleninu sodu obok siarki elementarnej nie podwyższał istotnie stężenia Se w surowicy, tak jak dodatek tylko seleninu. Także w porównaniu z kontrolą dodatek samej siarki nie zmieniał zawartości Se w wątrobie, ale istotnie obniżał jego wartość w mięśniach. Powyższe zmiany mogły być następstwem optymalnej zawartości Se w diecie przed doświadczeniem oraz wtórnej redystrybucji wewnątrzustrojowej i zwiększonego wydalania Se z moczem. Świadczy o tym istotnie niższe stężenie Se w mięśniach i nieco podwyższone w nerkach. Na możliwość zwiększonego wydalania Se z moczem przy nadmiarze S zwrócono uwagę jeszcze 50 lat temu (7), a nasilenie diurezy po dodatku siarki obserwowano także w naszym doświadczeniu. Obniżenie zawartości Se w mięśniach jest w jakimś stopniu potwierdzeniem udziału nadmiaru S w powstawaniu zaburzeń w metabolizmie Se i rozwoju objawów pokarmowej dystrofii mięśni.
Nadmiar S może też zmniejszać przyswajalność Ca i Mg i modyfikować ich przemianę oraz ograniczać efekty biologiczne również Fe i Zn (32), a także powodować zmiany i objawy charakterystyczne dla polioencefalomalacji – martwicy kory mózgowej, powstającej najczęściej na tle niedoboru witaminy B1. Polioencefalomalację na tle nadmiaru S obserwowano w warunkach naturalnych podczas żywienia paszą lub pojenia wodą o dużej zawartości związków S (14, 23, 30, 33), a także wywoływano doświadczalnie, stosując dietę bogatą w S (8, 27, 29). Objawy polioencefalomalacji wystąpiły m.in. u cieląt i jagniąt, gdy dla podwyższenia kwasowości diety w celu zapobiegania kamicy moczowej podawano siarczan amonu zamiast dwuwęglanu amonu (14).
Mechanizm niekorzystnego oddziaływania S na korę mózgową nie został dotychczas w pełni wyjaśniony. Objawom klinicznym i charakterystycznym zmianom w mózgu towarzyszyły różne stężenia witaminy B1 we krwi (8, 9, 27), co jednak nie wyklucza wpływu nadmiaru S na jej syntezę i metabolizm. Wykazano niedobór witaminy B1 u psów i kotów żywionych mięsem konserwowanym SO2 (33), ale także podwyższone jej stężenie we krwi u jagniąt z objawami polioencefalomalacji wywołanymi nadmiarem siarki w paszy (8). Być może nadmiar S oddziałuje na mózg zarówno za pośrednictwem witaminy B, jak i bezpośrednio, poprzez swoje toksyczne związki (23). W patogenezie zmian encefalomalatycznych na tle nadmiaru S uwzględnia się udział siarczynowych wolnych rodników, które wzmagając peroksydację lipidów, mogą uszkadzać tkankę nerwową (27). Siarczyny powstają u przeżuwaczy w żwaczu, a także w narządach w procesie oksydacji siarczków (26). Tkanka nerwowa zawiera dużo lipidów i wyjątkowo trudno i wolno ulega regeneracji.
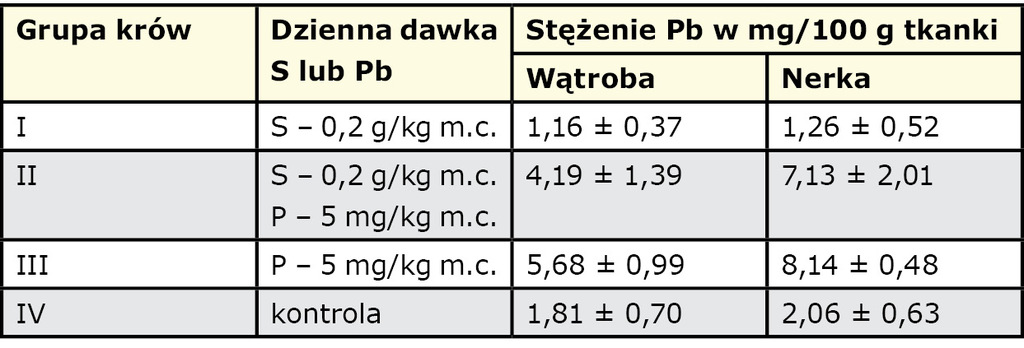
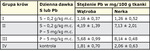
W pewnych okolicznościach nadmiar siarki w środowisku i w pokarmach może oddziaływać także pozytywnie. Jak wiadomo, siarka wchodzi w reakcje i wiąże nie tylko mikro- i makroelementy, ale także metale toksyczne, takie jak Pb, Cd i Hg (31). Powstające w wyniku tych reakcji siarczki i ich pochodne są dość trwałe, metabolicznie obojętne i zwykle wydalane z organizmu bez powodowania niekorzystnych zmian i zaburzeń. Proces wiązania i neutralizacji tych metali może mieć miejsce już w przewodzie pokarmowym, co utrudnia wchłanianie powstałych związków, a także w tkankach, gdzie pozostają, nie czyniąc większych szkód, albo są w mniejszym lub większym stopniu wydalane z moczem. Wydalanie nieorganicznych związków jest ułatwione diuretycznym oddziaływaniem siarki. Możliwość ograniczania gromadzenia się pierwiastków toksycznych w ważnych narządach poprzez dodatki siarki elementarnej do paszy potwierdzają wyniki badań przeprowadzonych na krowach, a dotyczących Pb (19).
Uzyskane wyniki ukazują wyraźną tendencję do zmniejszenia gromadzenia się ołowiu w wątrobie i w nerkach po 4-tygodniowym stosowaniu dodatku siarki elementarnej. Zawartość Pb była znacznie mniejsza u krów otrzymujących S w porównaniu z grupą kontrolną i mniejsza u krów otrzymujących Pb i S w porównaniu z grupą otrzymującą tylko Pb. Wzbogacenie diety w siarkę może podobnie oddziaływać na gromadzenie się Cd i Hg (11, 21) i ograniczać niekorzystne oddziaływanie nadmiernego, najczęściej przypadkowego podawania podstawowych mikroelementów. Z kolei niedobór S działa odwrotnie – zwykle wzmaga gromadzenie się metali toksycznych (31).
Mimo dużej emisji S do środowiska nadal występują jej niedobory w związku z intensyfikacją produkcji roślinnej i zwierzęcej. Szybki wzrost masy ciała zwierząt i wydajności produkcyjnej łączy się ze wzrostem zapotrzebowania na składniki mineralne pożywienia, w tym także na siarkę jako istotny makroelement warunkujący odpowiedni poziom produkcji i zachowanie dobrego stanu zdrowia. Niedobory siarki w roślinności i u zwierząt występują głównie na obszarach o ubogiej w składniki pokarmowe i przepuszczalnej glebie w czasie braku deszczu lub po małych opadach. W okresie suszy stężenie S w osoczu owiec spadało do 10 mg/L, ale nagle wzrastało do 20-40 mg/L w porze deszczowej (39). Spośród zwierząt niedobory S dotyczą przede wszystkim przeżuwaczy, u których zapotrzebowanie waha się w granicach 0,1-0,3% suchej masy paszy, średnio 0,18-0,20% suchej masy, ale może być znacznie wyższe w warunkach intensywnego wzrostu i tuczu oraz dużej produkcji mleka lub wełny (3). Stwierdzono m.in., że optymalne zapotrzebowanie na S elementarną u młodych buhajków wynosi około 0,3% suchej masy, ale znacznie wyższa zawartość siarki nie powodowała niekorzystnych zmian (5).
Wyraźne objawy niedoboru S u przeżuwaczy pojawiają się przy diecie zawierającej poniżej 0,1% S w suchej masie paszy. Zwierzęta na takiej diecie wykazują znaczne ograniczenie apetytu, spadek przyrostów masy ciała, wydajności mlecznej i przyrostu wełny. W zaawansowanej postaci widoczna jest osowiałość, osłabienie, wypadanie włosów i ślinienie, zdarzają się także zejścia śmiertelne (1, 37). Te zmiany wynikają m.in. ze zmniejszonej ilości i aktywności mikroflory żwacza oraz wtórnego niedoboru białka. Zmniejsza się wykorzystanie niebiałkowego N oraz synteza aminokwasów siarkowych i białka, a także biologicznie czynnych związków zapewniających prawidłowe funkcjonowanie organizmu (5). Do prawidłowego wykorzystania azotu niebiałkowego u przeżuwaczy stosunek S do N w paszy powinien być około 1 : 10, a nawet niższy u zwierząt wysoko produkcyjnych, szczególnie u owiec i kóz użytkowanych do produkcji wełny. Włosy i inne wytwory rogowe naskórka są wyjątkowo bogate w S (do 4%) i przy szybkim wzroście znacznie zwiększają zapotrzebowanie na ten pierwiastek. Dlatego wełniste owce i kozy są szczególnie wrażliwe na niedobór S.
Optymalny stosunek S do N dla roślin wynosi w granicach 1 : 20-35, a dla ludzi około 1 : 15 (59). Jak wspomniano, w stanach niedoboru S zmniejsza się przede wszystkim synteza aminokwasów siarkowych, a pośrednio białka, z czego wynikają różne zaburzenia metaboliczne. Istotny jest niedobór metioniny i cystyny, które tworzą strukturę białek. Metionina pełni m.in. rolę transportera grup metylowych, a cystyna jest produktem wyjściowym dla tak ważnych metabolicznie związków jak glutation i tauryna (4). Poza uczestnictwem w syntezie glutationu i w procesie metylacji, aminokwasy siarkowe są odpowiedzialne za syntezę białek ostrej fazy, uczestniczą w procesach oksydacyjno-antyoksydacyjnych i w odporności humoralnej, przez co warunkują wiele podstawowych funkcji komórek (2, 18, 36).
W stanach niedoboru siarki spada stężenie S w osoczu (< 10 mg/L) i metabolicznie czynnych związków zawierających siarkę, m.in. albumin, metioniny, glutationu (37), a także występuje tendencja do zwiększonego gromadzenia się w narządach pierwiastków toksycznych – Pb, Cd, Hg, i ważnych mikroelementów – Se, Cu, Zn – w przypadku nadmiernego ich podawania.
Wyniki naszych badań wskazują na częste występowanie niedoboru S, mimo regionalnie zwiększonej jej emisji do środowiska, a także różnorodność niekorzystnych konsekwencji niedoboru metabolicznie czynnych związków S dla stanu zdrowia zwierząt i ludzi. Dlatego S i jej związki są coraz częściej składnikiem nawozów mineralnych, pasz treściwych dla zwierząt i dodatków dietetycznych dla ludzi (37). Szczególnie cennym dodatkiem dietetycznym, a także uzupełniającym lekiem w niektórych zaburzeniach zdrowia są aminokwasy siarkowe (36).
Ryc. – T. Riha
PIŚMIENNICTWO
1. Ammerman C.B., Goodrich R.D.: Advances in mineral nutrition in ruminants. J. Anim. Sci. 57, 521-533, 1983. – 2. Atmaca G.: Antioxidant effects of sulfur-containing acids. Yonsci. Med. J. 45, 776-788, 2004. – 3. Bouchard R., Conrad H.R.: Sulfur metabolism and nutritional changes in lactating cows associated with supplemental sulfate and methionine analog. Can. J. Anim. Sci. 54, 587-593, 1974. – 4. Brosnan J.T., Brosnan M.E.: The sulfur-containing amino acids: a overiew. J. Nutr. 136, 1636s-1640s, 2006. – 5. Chalupa W., Oltjen R.R., Slyter L.L. et all.: Sulphur deficiency and tolerance in bull calves. J. Anim. Sci. 33, 278, 1971. – 6. Czauderna M.: Wpływ związków siarki i niektórych metali na poziom selenu w wybranych organach myszy i komórkach Saccharomyces cerevisiae. Rozpr. hab. Jabłonna 1999. – 7. Ganther H.E., Bauman C.A.: Selenium metabolism. II Modifying effects of sulfate. J. Nutr. 77, 408-414, 1962. – 8. Gooneratne S.R., Olkowski A.A., Christensen D.A.: Sulphur – induced polioencephalomalacia in sheep: some biochemical changes. Can. Vet. Rec. 53, 462, 1989. – 9. Goonerante S.R., Olkowski A.A., Klemmer R.G. et all.: High sulphur related thiamin deficiency in cattle. A field study. Can. Vet. J. 30, 139-146,1989. – 10. Grzebuła St.: Studia nad dystrofią pokarmową mięśni (PDM) u źrebiąt. Rozpr. hab. Lublin 1988.
11. El-Begearmi M.M., Ganther H.E., Sunde M.L.: Dietary interaction between methylmercury selenium, arsenic and sulfur amino acids in Japanese quail. Poultry Sci. 61, 272-279, 1982. – 12. Hard P.F., Ocumpaugh W.R., Greene L.W.: Forage mineral concentration, animal performance and mineral status of heifers cereal pastures fertilized with sulphur. J. Anim. Sci. G9, 2310-2320, 1991. – 13. Janowski T.M., Chmielowiec J.: Sulfuroza bydła. Medycyna Wet. 37, 265-268, 1981. – 14. Jeffrey M., Duff J.P., Higgiu R.J. et all.: Polioencephalomalacia associated with the ingestion of ammoniuom sulphate by sheep and cattle. Vet. Rec. 134, 343-348, 1994. – 15. Julian R.J., Harrison K.B.: Sulphur poisoning in cattle. Can. Vet. J. 16, 28-29, 1975. – 16. Kandylis K.: Toxicology of sulfur in ruminants: review. J. Dairy Sci. 67, 2179--2187, 1984. – 17. Kleczkowski M., Barej W., Kluciński W. et all.: Wpływ różnych stężeń Cu, Zn, Mo i S w paszy na zawartość Se w wątrobie buhajków. Zesz. Nauk PAN, 8, 136-141, 1994. – 18. Li P., Yin Y.L., Li D. et all.: Amino acid and immune function Br. J. Nutr. 98, 237-252, 2007. – 19. Madej E., Stec A., Filar J.: The influence of excess sulphur on selected blood indicators and lead accumulation in cow organs. Proceedengs XVIII World Buiatries Congress, 1373-1376. Bologna 1994. – 20. Madej E., Stec A., Filar J.: Wpływ nadmiaru siarki i jej związków na stan zdrowia zwierząt użytkowych. Materiały Sesji Naukowej KBN. Puławy 1995.
21. Madej E., Stec A., Riha T., Wieczorek J.: Wpływ dużej podaży siarki na stężenie metali ciężkich (Pb, Cd, Cu, Zn) w krwi, wątrobie i nerkach u krów. Materiały X Kongresu PTNW. Wrocław 1996. – 22. Mason J., Woods M., Poole D.B.R.: Accumulation of copper on albumin in bovine plasma in vivo after intravenous trithiomolybdate administration. Res. Vet. Sci. 41, 108-113, 1986. – 23. Mc Allister M.M., Gould D.H., Raisbeck M.F. et all:. Evaluation of ruminal sulfide concentrations and seasonal outbreaks of polioencephalomalacia in beet cattle in a feedlot. J. Am. Vet. Med. Assoc. 211, 1275-1279, 1997. – 24. Mills C.F., Dalgarno A.C., Bremner J., El-Galland T.T.: Influence of dietary content of molybdenium and sulphur upon hepatic retention of copper in young cattle. Proc. Nutr. Soc. 36, 105A, 1977. – 25. Moshtagni-Nia S.A., Derlin T.J., Phillips G.D.: Infuence of dietary copper, molybdenium and sulfur on copper metabolism of sheep. Can. J. Anim. 69, 187-194, 1989. – 26. Mottley C., Mason R.P.: Sulfate anion free radical formation by the peroxidation of (bi) sulfite and its reaction with hydroxyl radical scarengers. Arch. Biochem. Biophys. 267, 681-689, 1988. – 27. Olkowski A.A., Goonerante S.R., Rousseaux C.G., Christensen D.A.: Role of thiamine status in sulphur induced polioencephalomalacia in sheep. Res. Vet. Sci. 52, 78-85, 1997. – 28. Riha T.: Wpływ zwiększonej podaży siarki na metabolizm selenu z uwzględnieniem stanu antyoksydacyjnego u buhajków. Rozpr. dokt. Lublin, 2001. – 29. Rousseau C.G., Olkowski A.A., Chauvet A. et all.: Ovine polioencephalomalacia associated with dietary sulphur intake. J. Vet. Med. A. 38, 229, 1991. – 30. Smart M.E., Cohen R., Christensen D.A., Coilliams C.M.: The effects of sulphate removal from the drinking water on the plasma and liver cooper and zinc concentrations of beet cows and their calves. Can. J. Amin. Sci. 66, 669, 1986.
31. Smith G.M., White C.L.: A molybdenum – sulphur – cadmium interaction in sheep. Aust. J. Agric. Res. 48, 147-154, 1997. – 32. Spears J.W., Burns J.C., Hatch P.A.: Sulfur fertilization of cool season grasses and effect on utilization of minerals, nitrogen and fiber by steers. J. Dairy Sci. 68, 347-355, 1985. – 33. Studdert V.P., Labue R.H.: Thiumin deficiency in cats and dogs associated with feeding meat preserved with sulphur dioxide. Aust. Vet. J. 68, 54-56, 1991. – 34. Suttle N.F.: Effects of organic and inorganic sulphur on the availability of dietary copper to sheep. Br. J. Nutr. 32, 559-568, 1974. – 35. Suttle N.F.: The interactions between cooper molybdenum and sulphur in ruminant nutrition. Anna. Rev. Nutr. 11, 121-140, 1991. – 36. Tesserand S., Metayer Constard S., Collin A. et all.: Role of sulfur aminoacids in controlling nutrient metabolism all functions: implications for nutrition. Br. J. Nutr. 10, 1132-1139, 2008. – 37. Underwood E.J., Suttle N.F.: Mineral Nutrition of Livestock. 231-250, CAB International 1999. – 38. White J.B.: Sulphur poisoning in ewes. Vet. Rec. 76, 279-280, 1964. – 39. White C.L., Kumagai H., Barnes M.: The sulphur and selenium status of pregnant ewes grazing Mediterranean pastures. Aust. J. Agric. Res. 48, 1081-1087, 1997. – 40. Wieczorek J.: Wpływ nadmiernej podaży siarki na metabolizm miedzi u buhajków. Rozpr. dokt. Lublin 2000.